
El propósito de la respiración celular es convertir la energía almacenada en la glucosa en trifosfato de adenosina (ATP), la principal moneda energética de la célula. La respiración celular ocurre en tres etapas principales. La glucólisis implica la descomposición de la glucosa en piruvato, produciendo una pequeña cantidad de ATP. El ciclo del ácido cítrico descompone aún más el piruvato en dióxido de carbono, generando NADH y FADH2. El paso final de la respiración celular es la cadena de transporte de electrones y la fosforilación oxidativa, que producen una gran cantidad de ATP, así como agua como subproducto. En este artículo, analizaré las características de la vía glucolítica que resisten la explicación mediante procesos evolutivos no dirigidos y que proporcionan evidencia de un Diseño Inteligente.
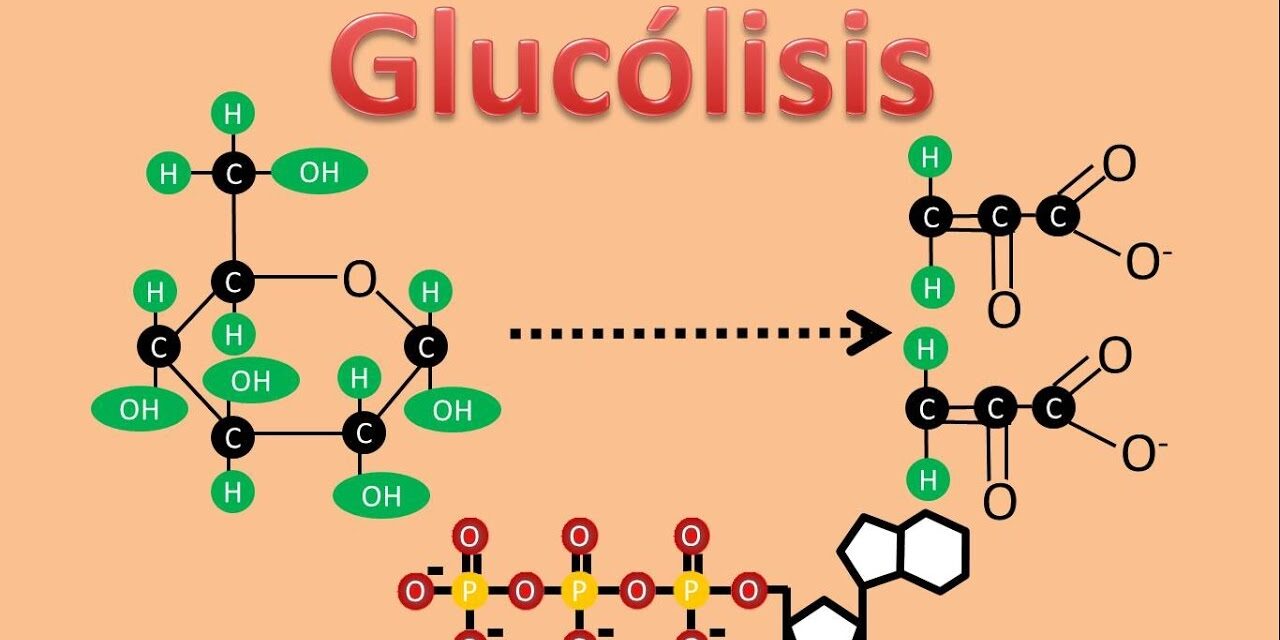
El primer paso en la representación celular (glucólisis) está representado por la figura de la parte superior. La glucólisis es omnipresente en todos los organismos vivos. Como se muestra en la figura, la glucólisis implica la conversión de glucosa, a través de una serie de intermediarios, en piruvato. Este piruvato es luego transportado a las mitocondrias, donde la enzima piruvato deshidrogenasa lo convierte en acetil-CoA. Este proceso también produce NADH y libera una molécula de dióxido de carbono (CO2). El acetil-CoA luego alimenta el ciclo del ácido cítrico, donde se oxida aún más, generando más NADH, FADH2 y ATP (o GTP).
¿Evolución incremental?
Se ha propuesto que la glucólisis es la primera vía bioquímica que surgió en la evolución. Entre las razones para ello se encuentra el hecho de que la glucólisis se encuentra en todas partes en el árbol de la vida (por lo que se puede inferir que estuvo presente en el último ancestro común universal). Además, la glucólisis es una secuencia de reacción anaeróbica y, por lo tanto, es coherente con la ausencia de oxígeno en el entorno primitivo de la Tierra.
Sin embargo, existen desafíos significativos para el propuesto origen evolutivo de la vía de la glucólisis. Por ejemplo, la conversión de glucosa en piruvato involucra hasta diez enzimas independientes, típicamente de 300 a 500 aminoácidos de longitud. Es extremadamente improbable que diez enzimas con actividades complementarias pudieran haber surgido esencialmente al mismo tiempo. Pero ¿podría esta vía haber evolucionado de manera incremental, ya sea hacia adelante o hacia atrás? En general, se rechaza que la glucólisis haya surgido de manera inversa (es decir, que inicialmente se dispusiera del piruvato, luego de su precursor, etc.), ya que no era el piruvato oxidado, sino el azúcar, lo que habría estado presente en el entorno de la Tierra primitiva. Además, cada intermediario entre la glucosa y el piruvato está fosforilado (es decir, uno o dos de sus grupos hidroxilo son reemplazados por fosfato). Esto implica una reacción de condensación (en la que se elimina una molécula de agua). Dadas las dificultades de este tipo de reacción, es cuestionable si los diversos intermediarios podrían haber surgido de manera abiótica en cantidades suficientemente altas para facilitar el origen de la glucólisis.
La opinión más popular es que la glucólisis evolucionó de manera incremental en la dirección hacia adelante. Esta hipótesis, por supuesto, se basa en la presunción de que los intermediarios podrían haber tenido su propia utilidad independiente. Sin embargo, dado que generalmente se piensa que la glucólisis surgió muy temprano (antes de que pudiera haber surgido una utilidad adicional de los intermediarios), parece improbable que los intermediarios pudieran haber tenido usos independientes.
Causa circular
Obsérvese que el proceso de glucólisis consume dos moléculas de ATP ─una en el paso de glucosa a glucosa 6-fosfato (catalizada por la hexoquinasa) y otra en el paso de fructosa 6-fosfato a fructosa 1,6-bisfosfato (catalizada por la fosfofructoquinasa). El rendimiento total de ATP de la glucólisis es cuatro (aunque se producirán muchos más ATP más adelante), mientras que se consumen dos, lo que hace que el rendimiento neto sea dos ATP. Para que se produzca ATP, primero debe consumirse ATP. Esto presenta un desafío de circularidad causal para una explicación evolutiva de los orígenes de la glucólisis. Sorprendentemente, esta circularidad causal de ATP necesario para fabricar más ATP parece ser omnipresente a lo largo de la vida1. ¿Cómo podría establecerse el proceso de glucólisis sin un suministro inicial de ATP? Además, después del consumo del primer ATP, hay al menos cinco pasos adicionales (cada uno con su propia enzima) antes de que se produzca más ATP, y nueve antes de que haya un rendimiento neto de ATP. Dado que la selección natural carece de previsión, esto hace que sea extremadamente improbable que las enzimas iniciales de la vía glucolítica pudieran haber servido para algún beneficio en ausencia de las enzimas posteriores en la vía.
Excluyendo el agua
De las diez enzimas que intervienen en la glucólisis, seis catalizan reacciones que implican la transferencia de un grupo fosfato. Que un fosfato reaccione con un grupo hidroxilo del agua para formar ácido fosfórico es tan favorable energéticamente como que reaccione con el hidroxilo, un azúcar o ADP, pero esto no supondría ninguna ventaja evolutiva. Por tanto, el agua debe excluirse de los sitios activos de las enzimas para evitar que se produzcan reacciones hidrolíticas. Esto se consigue mediante un mecanismo que implica cambios conformacionales que se asemejan a un «movimiento de bisagra». Inicialmente, el sitio activo de la enzima asume una conformación abierta, lo que permite la entrada del sustrato. Cuando el sustrato se une al sitio activo, induce un cambio conformacional que hace que la enzima experimente un movimiento de «cierre», en el que los dominios de la enzima se unen, protegiendo eficazmente el sitio activo. Este movimiento no sólo asegura el sustrato, sino que también impide que el agua entre en el sitio activo.
Este fenómeno subraya la sofisticación de la ingeniería (y el grado de especificidad de los aminoácidos) de estas enzimas. Dado que la exclusión del agua es absolutamente crítica para que se produzcan las reacciones adecuadas, no tendría sentido tener una enzima parcialmente formada (es decir, una que pudiera catalizar la reacción de fosforilación pero que no lograra excluir el agua). Esto genera más dudas sobre la capacidad de las adaptaciones incrementales para explicar la vía glucolítica.
¿Relación entre enzimas?
Otra cuestión es que, si la glucólisis fuese una de las primeras vías metabólicas que evolucionó, cabría esperar que en el momento de su origen sólo existiera un pequeño repertorio de enzimas. Además, los compuestos sobre los que actúan estas enzimas tienen estructuras similares. Esto podría llevarnos a predecir que las enzimas implicadas en la glucólisis están relacionadas evolutivamente entre sí. Sin embargo, como señala Keith Webster:
Los datos secuenciales y cristalográficos favorecen la evolución divergente de, por ejemplo, la monofosfoglicerato mutasa y la difosfoglicerato mutasa, y posiblemente la gliceraldehído-3-P deshidrogenasa y la fosfoglicerato quinasa a partir de respectivos ancestros comunes, pero la convergencia parece haber desempeñado un papel más importante en el desarrollo de las otras 11 enzimas (Fothergill-Gilmore, 1986; Fothergill-Gilmore y Watson, 1989). Por ejemplo, no hay evidencia de un ancestro común para ninguna de las cuatro quinasas glucolíticas ni de las siete enzimas que se unen a los nucleótidos, con la excepción de las mencionadas anteriormente. Más bien, parece probable que la vía fuera el resultado del ensamblaje casual de enzimas y genes que evolucionaron independientemente, probablemente en asociación con la coevolución de otras funciones y vías vinculadas. [Énfasis añadido.]2
Esto parece contrario a lo que podría predecirse a partir de una explicación evolutiva de los orígenes de la glucólisis.
Diseño Inteligente
La explicación de la vía glucolítica en términos de mecanismos evolutivos no dirigidos plantea múltiples desafíos. La complejidad y la sofisticación de la ingeniería se ajustan mucho mejor a la hipótesis del Diseño. En particular, la circularidad causal del ATP necesario para producir más ATP es difícil de explicar mediante un proceso evolutivo gradual. Por otra parte, este tipo de fenómeno no resulta en absoluto sorprendente si se supone que interviene una mente inteligente.
Notas
- Kun A, Papp B, Szathmáry E. Computational identification of obligatorily autocatalytic replicators embedded in metabolic networks. Genome Biol. 2008;9(3): R51.
- Webster KA. Evolution of the coordinate regulation of glycolytic enzyme genes by hypoxia. J Exp Biol. 2003 Sep; 206(Pt 17):2911-22.
Artículo publicado originalmente en inglés por Jonathan McLatchie en Evolution News & Science Today




{kind=link}