
¿Hay rutas largas, graduales, de estructuras intermedias funcionales, separadas por una o quizás algunas mutaciones, que conducen a cada especie y a cada diseño y estructura en toda la biología? Como vimos aquí, este ha sido un reclamo fundamental y una expectativa de la teoría de la evolución que está en desacuerdo con la ciencia. Algunos evolucionistas han intentado evitar el problema con la teoría neutral, pero eso solo empeora las cosas.
Si una mutación es rara, muchas mutaciones son astronómicamente raras. Por ejemplo, si una mutación en particular tiene una probabilidad de uno en cien millones (1 en 108) de ocurrir en una nueva persona, entonces cien mutaciones particulares de ese tipo tienen una probabilidad de uno en 10800 de ocurrir. No va a suceder. Echemos un vistazo a un ejemplo: células nerviosas y sus señales de potencial de acción.
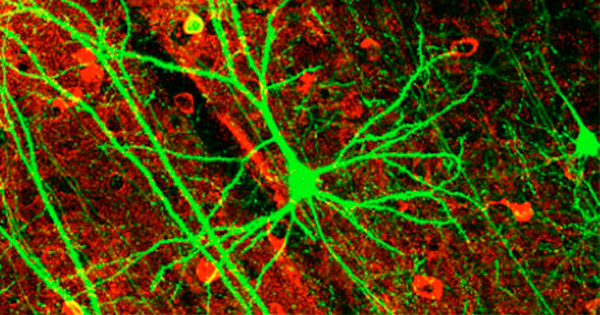
Las células nerviosas tienen una cola larga que lleva un impulso electrónico. La cola puede tener varios pies de largo y su señal puede estimular la acción de un músculo, controlar una glándula o informar una sensación al cerebro.
Como un cable que contiene miles de cables telefónicos diferentes, las células nerviosas a menudo se agrupan para formar un nervio. Los primeros investigadores consideraron que tal vez el impulso electrónico viajó a lo largo de la cola de la célula nerviosa como la electricidad en un cable. Pero pronto se dieron cuenta de que la señal en las células nerviosas es demasiado débil para viajar muy lejos. La célula nerviosa necesitaría aumentar la señal a lo largo del camino para que viaje a lo largo de la cola.
Después de años de investigación, se descubrió que la señal está impulsada por las proteínas de la membrana. En primer lugar, hay una proteína de la membrana que bombea simultáneamente dos iones de potasio a la célula y tres iones de sodio fuera de la célula. Esto establece un gradiente químico a través de la membrana. Hay más potasio dentro de la célula que afuera, y hay más sodio afuera que adentro. Además, hay más iones cargados negativamente dentro de la célula por lo que hay una caída de voltaje (50-100 milivoltios) a través de la membrana.
Además de la bombeo de sodio y potasio, también hay canales de sodio y potasio. Estas proteínas de membrana permiten que el sodio y el potasio, respectivamente, pasen a través de la membrana. Normalmente están cerrados, pero cuando el impulso electrónico en descomposición viaja a lo largo de la cola de la célula nerviosa, hace que los canales de sodio se abran rápidamente. Los iones de sodio que están fuera de la célula luego ingresan a la célula por el gradiente electro-químico. Como resultado, la caída de voltaje se invierte y el impulso electrónico en descomposición, que provocó que se abran los canales de sodio, aumenta a medida que avanza a lo largo de la cola de la célula nerviosa.
Cuando el voltaje va de negativo a positivo dentro de la célula, los canales de sodio se cierran lentamente y los canales de potasio se abren. Por lo tanto, los canales de sodio están abiertos solo momentáneamente, y ahora con los canales de potasio abiertos, los iones de potasio concentrados dentro de la célula salen por su gradiente electroquímico. Como resultado, se restablece la caída de voltaje original.
Este proceso se repite a medida que el impulso electrónico viaja a lo largo de la cola de la célula nerviosa, hasta que el impulso finalmente llega al final de la célula nerviosa. Aunque hemos omitido muchos detalles, debería ser obvio que el proceso depende del funcionamiento intrincado de las tres proteínas de la membrana. La bomba de sodio y potasio ayuda a configurar el gradiente petroleoquímico, el impulso electrónico es lo suficientemente fuerte como para activar el canal de sodio, y luego los canales de sodio y potasio se abren y cierran con una sincronización precisa.
¿Cómo, por ejemplo, los canales diseñados para ser selectivos de iones? El sodio es aproximadamente 40 por ciento más pequeño que el potasio, por lo que el canal de sodio puede excluir el potasio si es lo suficientemente grande para el sodio. Las mutaciones aleatorias deben haber golpeado en una secuencia de aminoácidos que se doblaría justo para proporcionar el tamaño del canal correcto.
El canal de potasio, por otro lado es lo suficientemente grande para el potasio y el sodio, sin embargo, es altamente eficiente. De alguna manera excluye el sodio casi a la perfección (la relación de potasio a sodio es de aproximadamente 10000), pero permite que pase el potasio casi como si no hubiera nada en el camino.
Las células nerviosas se disparan constantemente en su cuerpo. Controlan tus ojos mientras lees estas palabras, y envían al cerebro las imágenes que ves en esta página. Ellos, junto con las señales químicas, controlan una multitud de procesos en nuestros cuerpos, y no hay ninguna razón científica para pensar que evolucionaron gradualmente, una mutación a la vez.
De hecho, esa idea contradice todo lo que sabemos de la ciencia. Y, sin embargo, esto es lo que los evolucionistas creen. Permítanme repetirlo: los evolucionistas creen que las células nerviosas y sus diseños de potencial de acción evolucionaron una mutación a la vez. De hecho, los evolucionistas creen que esto es un hecho comprobado, más allá de toda duda razonable.
Sería difícil imaginar una afirmación más absurda. Así que echemos un vistazo a los detalles de esta línea de pensamiento. Aquí hay un documento reciente de la Royal Society, que representa el estado del arte en el pensamiento evolutivo sobre este tema. El documento pretende proporcionar una explicación detallada de cómo la evolución temprana produjo la tecnología del potencial de acción.
Suena prometedor, pero cuando los evolucionistas hablan de «detalles», tienen algo ligeramente diferente en mente. Aquí hay varios pasajes del documento que revelan que no solo hay una falta de detalles, sino que el estudio no es completamente científico.
Proponemos que el siguiente paso en la evolución de DCS eucariota [acoplamiento de la despolarización de la membrana (a través de la afluencia incontrolada de calcio), la contracción y la secreción] ha sido el reclutamiento de canales de calcio sensibles al estiramiento, que permiten la afluencia controlada de calcio sobre el estrés mecánico antes de que el daño real se produzca, y así anticipar los efectos de la rotura de la membrana. [Énfasis añadido.]
¿El reclutamiento de canales de calcio? ¿Y exactamente quién hizo el reclutamiento? Aquí los autores confían en la vaga terminología para escribir sobre una serie de detalles problemáticos de cómo las mutaciones aleatorias de alguna manera realizaron este reclutamiento.
Para prevenir la ruptura real, el primer papel de los canales de Ca ++ mecanosensoriales podría haber sido activar previamente los componentes de la vía de reparación en las membranas estiradas.
¿»Para prevenir»? Vamos a deletrear la lógica un poco más claro. Los autores están formulando la hipótesis de que estos canales de calcio desarrollaron la capacidad de activar previamente la vía de reparación para «prevenir» la ruptura real. Al deletrear la lógica un poco más claramente, podemos ver más fácilmente la teleología habitual en la investigación. La literatura de la evolución está llena de teleología, y por buenas razones. Los evolucionistas no pueden formular y expresar sus ideas sin ella. El aristotelismo está muerto – viva el aristotelismo.
Como otro paso anticipado, la actomiosina podría haberse colocado previamente debajo de la membrana plasmática (de ahí la red de actomiosina cortical detectada en cada célula eucariótica) y también podría haber evolucionado la sensibilidad directa al estiramiento… Una vez adquirida su posición cortical y su mecanosensibilidad, la red de actomiosina podría cumplir automáticamente una función adicional: mantenimiento de la forma de la celda, ya que cualquier deformación celular localizada estiraría la red de actomiosina cortical y desencadenaría una contracción compensatoria inmediata. Esta propiedad habría surgido como un efecto secundario (un «spandrel») de la presencia de actomiosina cortical para la reparación de la membrana, y rápidamente resultó ventajosa.
¿Un «paso anticipado»? «Preposicionamiento»? La actomiosina «evolucionó» la sensibilidad al estiramiento? La posición y la mecanosensibilidad «fueron adquiridas»? La red podría «cumplir una función adicional»? Lo sentimos, pero las máquinas moleculares (como la actomiosina) no «evolucionan» nada. Hay más teleología empaquetada en estas pocas oraciones que cualquier tratado medieval. Y para una buena medida, los autores también agregan la asombrosa casualidad de que esta función adicional «habría surgido como un efecto secundario». Eso fue una suerte.
Una vez que cubría la corteza celular, la red de actomiosina adquirió la capacidad de deformar la célula mediante contracción localizada.
¿La red de actomiosina «adquirió la capacidad» de deformar la célula mediante contracción localizada? Movimiento inteligente por parte de la red. ¿Pero podemos preguntar cómo sucedió eso?
Basado en el estudio genómico del protista Naegleria que tiene un ciclo de vida bifásico (alternando entre una fase ameboide y una fase flagelada), se ha propuesto que la locomoción ameboide es ancestral para los eucariotas. Puede haber evolucionado en ambientes intersticiales confinados, ya que es particularmente instrumental para las células que necesitan moverse a través de espacios pequeños de forma irregular mediante deformación exploratoria.
La locomoción ameboide evolucionó «ya que es particularmente instrumental». No tiene hace uso del infinitivo, pero esto no es menos teleológico. Las cosas no evolucionan porque son «instrumentales». Lo que los autores no informan a sus lectores es que esto requeriría un enorme número de mutaciones aleatorias.
Uno puede hipotetizar que, si los canales de calcio sensibles al estiramiento y la actomiosina cortical formaran parte del juego de herramientas ancestrales eucariotas (como lo indica la genómica comparativa), la deformación de la membrana en un entorno confinado probablemente desencadenaría la afluencia de calcio al abrir canales sensibles al estiramiento, lo que a su vez inducen una contracción amplia de la actomiosina a través de la parte deformada de la corteza celular, la deformación global y el movimiento celular lejos de la fuente de presión.
El concepto de un «juego de herramientas moleculares» es estándar en el pensamiento evolutivo y otro ejemplo de pensamiento teleológico.
Por lo tanto, se puede proponer que una forma ancestral simple de movimiento ameboide evolucionó como una consecuencia natural del escenario descrito anteriormente para el origen de la actomiosina cortical y el acoplamiento de contracción del calcio; una vez establecido, se pudo volver más complejo.
El movimiento ameboide evolucionó «como una consecuencia natural» y «una vez establecido» se volvió «más complejo». Esto no es más que una narración teleológica sin evidencia de respaldo.
Por lo tanto, es tentador especular que, una vez que la señalización de calcio había ganado control sobre las formas primitivas de movimiento ameboide, el mismo sistema de señalización comenzó a modificar el latido ciliar, posiblemente para «cambiar» entre estados locomotores.
¿La señalización de calcio «ganó control» y luego «comenzó a modificar» la vibración ciliar «para» cambiar «entre estados locomotores»? El «para cambiar» es otra expresión que recuerda a la teleología, y «control ganado» es un movimiento activo del sistema de señalización de calcio. Teleología pura, no adulterada.
Posiblemente, en los eucariotas ancestrales el calcio indujo un cambio relativamente simple (como el paro ciliar, como se observa en muchas células animales y en Chlamydomonas en respuesta a altas concentraciones de Ca ++), que luego se modificó gradualmente en modulaciones más sutiles del modo de latido con un ayuno volumen de negocios de los actores moleculares mediado por la adición diferencial, la complementación y la pérdida.
¿»El calcio indujo un cambio relativamente simple»? Lo sentimos, los iones no inducen interruptores, simples o de otra manera. Y el interruptor ¿»luego se modificó gradualmente en modulaciones más sutiles»? Tenga en cuenta cómo la voz pasiva obvia esos detalles espinosos. El modificador «se modificó» omite convenientemente el hecho de que dicha modificación tendría que producirse mediante una mutación aleatoria, una mutación a la vez.
Alternativamente, el control de los cilios por el calcio podría haber evolucionado de manera convergente, pero tal convergencia habría sido extraordinariamente omnipresente, ya que no parece existir un flagelo eucariótico que no esté controlado por el calcio de una forma u otra.
¿»Podría haber evolucionado de manera convergente»? ¿Y exactamente cómo sucedería eso? Al menos los autores luego admiten lo absurdo de esa alternativa.
Lamentablemente, carecen de esa sensibilidad para el resto del documento. Como vimos anteriormente, el documento se basa en una secuencia de pensamiento teleológico. Entra en el género evolutivo donde la evolución se toma, a priori, como algo dado. Esta suposición respalda vastas extensiones de pensamiento teleológico y narración de cuentos a nivel de historieta. No solo hay una falta de apoyo empírico, sino que el género es totalmente no científico, como lo revela incluso una lectura medianamente crítica.
Y no hace falta decir que el documento no hace absolutamente nada para aliviar el problema con el que comenzamos. Los muchos saltos de lógica y razonamiento en el documento revelan todo tipo de cambios monumentales que la evolución requiere para construir células nerviosas y la tecnología del potencial de acción. No estamos viendo una narración de cambios pequeños y graduales, cada uno contribuyendo a la aptitud general. Muchas, muchas mutaciones simultáneas serán necesarias. Incluso un número mínimo conservador de 100 mutaciones simultáneas conduce al resultado insostenible de una probabilidad de 1 en 108 de ocurrir.
Simplemente no va a suceder.




{kind=link}