
1. Introducción: ¿Por qué una distancia biológica que es actualmente imposible de medir, profundidad ontogenética, sin embargo realmente importa a la teoría evolutiva?
La teoría de la selección natural proporciona una explicación mecanicista y causal de cómo los seres vivos llegaron a verse como si hubieran sido diseñados para un propósito.
– Richard Dawkins (1982, 45)La selección natural no nos dice absolutamente nada sobre los mecanismos subyacentes de los cambios genómicos, o sus consecuencias sobre los cambios de desarrollo que conducen a innovaciones evolutivas. En resumen, no dice nada sobre la aparición de formas complejas.
– George L.G. Miklos (1993, 24)
¿Cómo puede una teoría de la evolución que pretende explicar cómo las criaturas con trillones de células surgieron de los comienzos multicelulares se toman en serio si todo lo que nos dice es que las tasas de destrucción pueden alterar la composición genética de las poblaciones?
– Wallace Arthur (2004, 36)
Una conclusión sólida es que el proceso evolutivo que generó los programas genómicos responsables de la formulación evolutiva de los planes corporales eumetazoarios básicos fue en muchos aspectos muy diferente de los cambios evolutivos que se pueden observar a nivel de especie en los animales modernos.
– Eric Davidson y Douglas Erwin (2010, 1)
Esas declaraciones te dicen dónde va esta serie de blogs de profundidad ontogenética (PO). Aquí está mi tesis: la teoría de la evolución por selección natural no explica el origen de la forma animal, porque la selección natural no puede explicar el origen de novo de las etapas de desarrollo requeridas para construir animales (es decir, evolucionar). El concepto de PO nos ayuda a entender por qué.
Para empezar, quiero mostrar por qué PO es fácil de entender, en cierto sentido, de todos modos, pero, por el momento, casi imposible de medir. Para que la macroevolución de los animales se produzca de nuevo a través de la selección natural, debe atravesarse la profundidad ontogenética. Este es un problema ineludible, y actualmente no resuelto. Y cada mejora en la medición de PO solo hará que el problema sea más desafiante.
2. Una distinción útil: denominación versus medición
En este momento, los cielos son claros en Chicago. Mirando hacia el cielo del sudeste, puedo ver el sol, a cierta distancia en el espacio.
Ahora, supongamos que estamos mirando al sol hace 500 años, antes que Kepler, Newton, Huygens y Cassini. Sabemos que el sol está a una distancia mensurable de la Tierra, y podemos darle un nombre a esa distancia. Llámalo la distancia Tierra-Sol, o, como ahora se conoce, la Unidad Astronómica (UA, ver Figura 1).

Figura 1
Entonces, ¿qué tan lejos está exactamente el Sol de la Tierra? Hace quinientos años, no teníamos telescopios, radares, satélites ni ninguno de los otros instrumentos actualmente utilizados para medir la UA. Pero sí teníamos geometría, y podíamos captar el concepto de paralaje. Debería ser bastante fácil, usando observaciones a simple vista del Sol y los planetas, estimar el valor exacto de la UA.
Realmente no. Como muestra la historia de la astronomía, las mediciones (o estimaciones) de la UA, intentadas desde la antigüedad, no se asentaron en nada cercano al valor moderno hasta finales del siglo XVIII. Los datos requeridos para realizar la triangulación representada en la Figura 2, con precisión, empleando el Tránsito de Venus, simplemente no estaban disponibles, o se sabía que eran demasiado imprecisos para ser útiles.
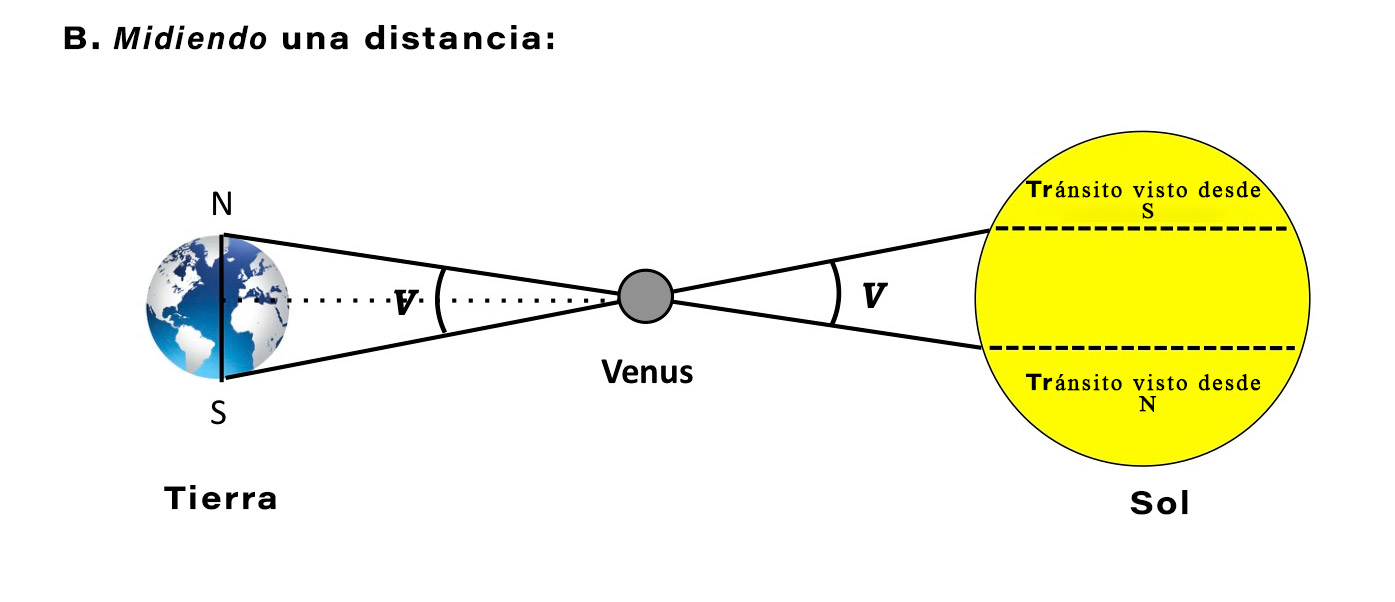
Figura 2
¿Eso significa que el concepto de UA es ininteligible? Por supuesto que no. Las dificultades que enfrentamos al medir una distancia no implican que la distancia en sí sea irreal o sin importancia. Las dificultades solo significan que la distancia puede ser difícil de especificar en términos exactos.
3. Profundidad ontogenética: denominación versus medición
Transicionando a la profundidad ontogenética. Aquí está mi formulación original (2003, 459):
La profundidad ontogenética estima la distancia, en términos de división y diferenciación celular, entre una condición unicelular y el metazoo adulto macroscópico capaz de reproducirse.
Las palabras en negrita definen, o nombran, una distancia. Las palabras en cursiva, por el contrario, representan un intento para medir esa distancia. Si el lector puede captar esta distinción, es decir, entre definir o nombrar una distancia, versus medirla, gran parte de la confusión que rodea la profundidad ontogenética se disuelve. El punto importante para recordar es esto: la distancia de (PO) es real, y debe ser atravesada por cualquier proceso evolutivo candidato, ya sea que podamos medir la PO con precisión o no.
Considere un paralelismo para definir versus medir la UA. La Figura 3 muestra la PO como la distancia entre un huevo fertilizado de C. elegans y el fenotipo adulto capaz de reproducirse. Podemos pensar en esto como mirar el Sol y saber que está a cierta distancia [intervalo finito] en el espacio.
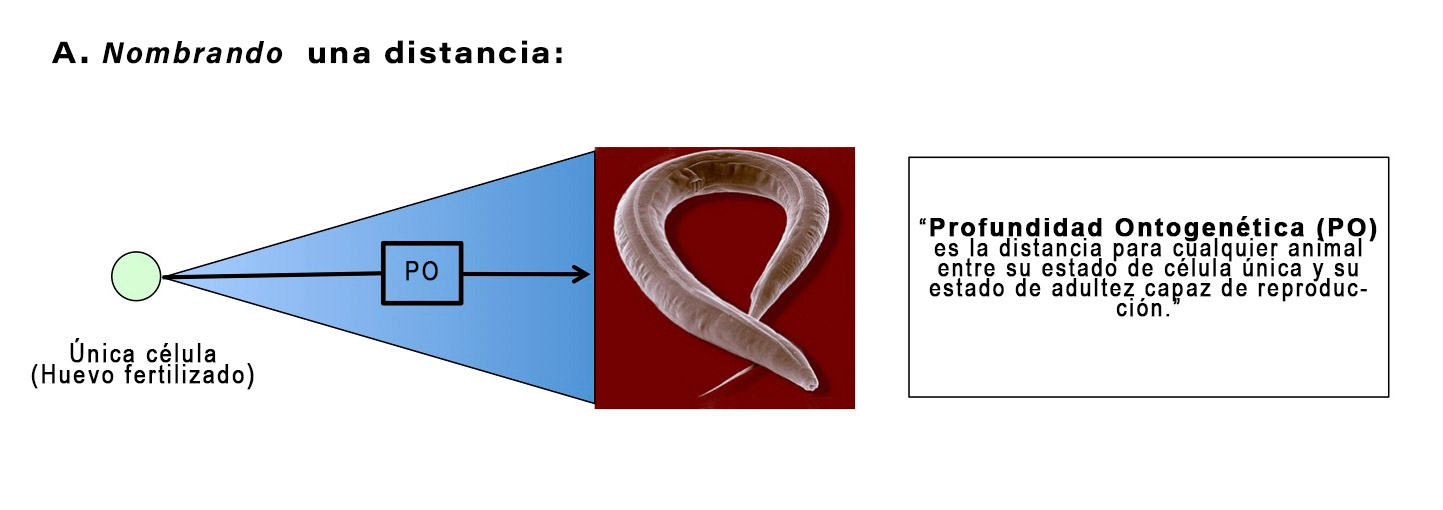
Figura 3
La figura 4, por el contrario, postula que la división y diferenciación celular (es decir, tipos de células) proporcionan las unidades correctas para medir, es decir, dar un valor exacto, a la PO:
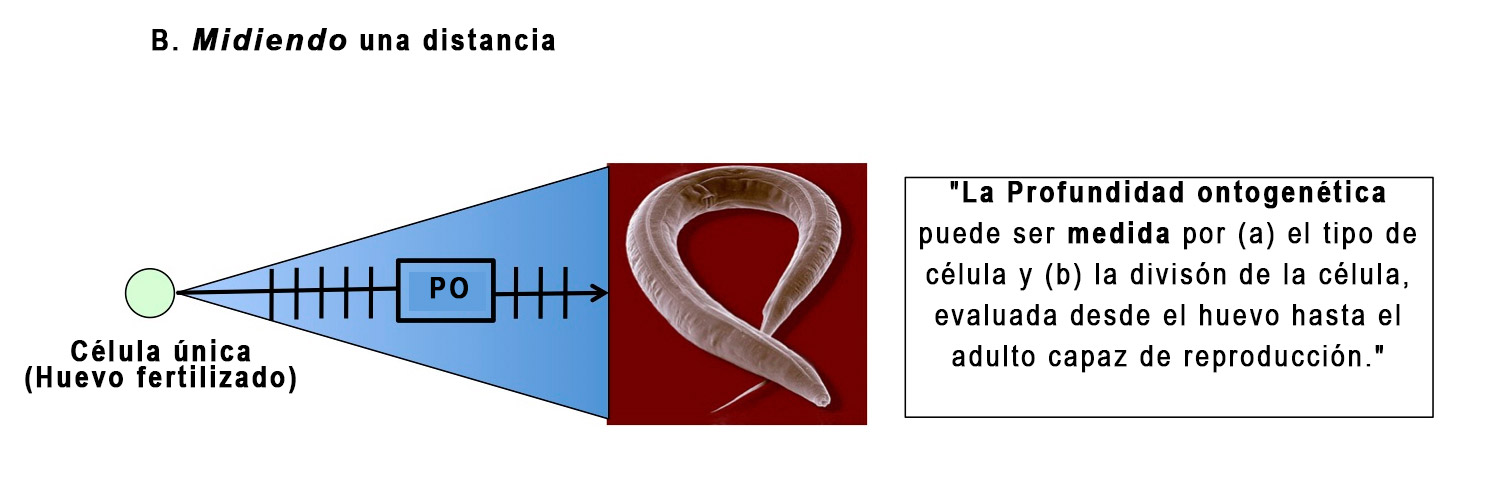
Figura 3
¿Ve la diferencia? Cualquiera estaría de acuerdo en que hay una distancia entre el huevo y el adulto en C. elegans. La pregunta es cuál es la mejor manera de medir esa distancia, en unidades (o medidas) que permiten la comparación con otros grupos de animales. En su crítica original, Myers dio algunas razones para pensar que las métricas del tipo de célula y la división celular no funcionarían. Pero él subestimó la dificultad, como lo ilustraré a continuación.
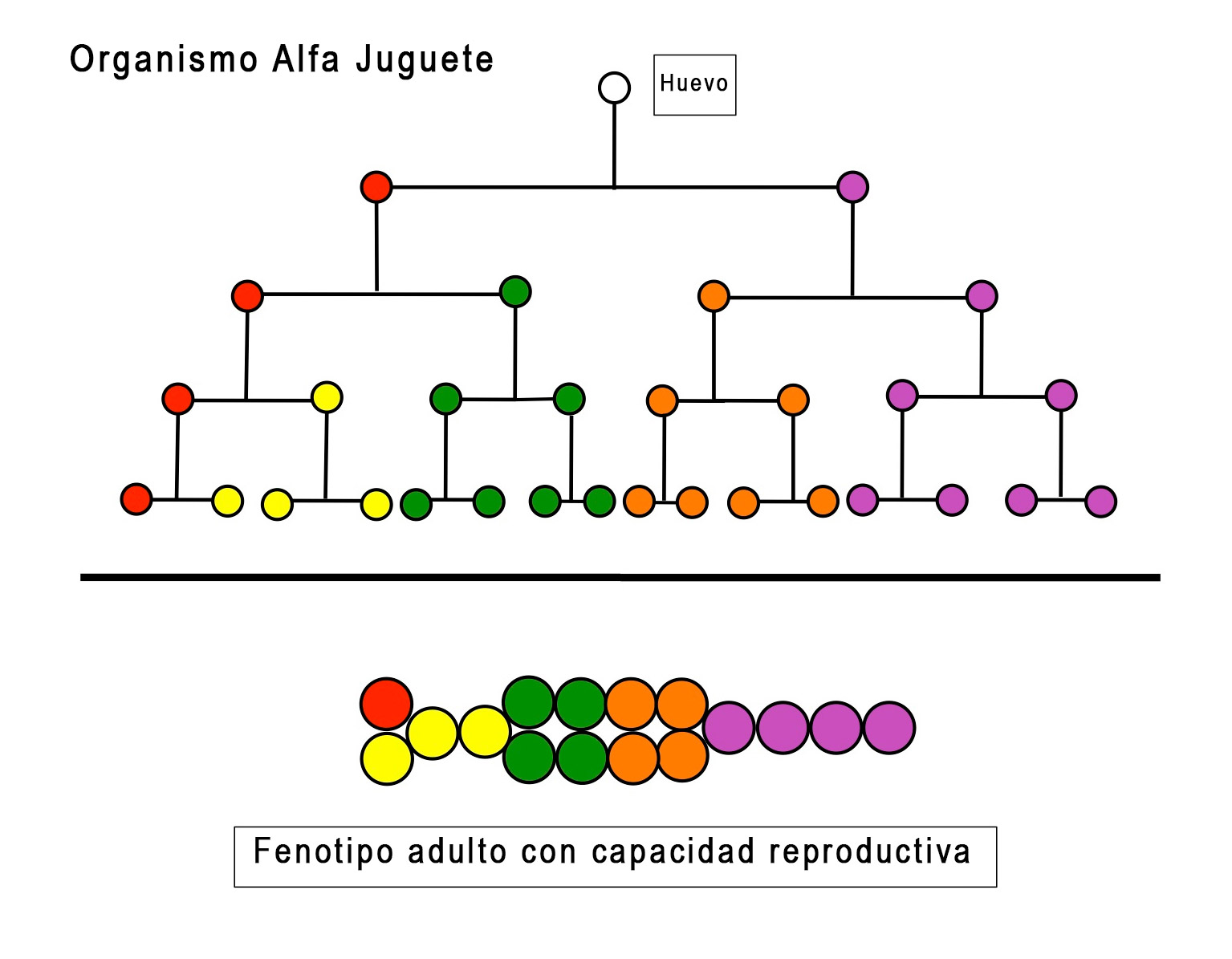
Aquí está el linaje celular (ver Figura 5) de un organismo alfa juguete, con 16 celdas de 5 tipos. Cuatro rondas de división celular separan cualquier célula en el punto de partida adulto del huevo fertilizado. Una medida simple de PO para este organismo de juguete multiplicaría el número promedio de divisiones celulares, entre el huevo y cualquier célula en el adulto (4), multiplicado por el número total de tipos de células (5, con el glóbulo rojo como línea germinal produciendo gametos), para producir una PO de 20.

Figura 5
Pero compare el organismo juguete alfa con el organismo juguete beta. Si bien tienen la misma PO, si usamos la métrica simple de 4 rondas de división celular X 5 tipos de células = 20, sus formas adultas respectivas son muy diferentes. Beta tiene una gran cavidad interna, y coloca su «gónada» (la célula roja) dentro de la pared de esa cavidad, mientras que la gónada de alfa es exterior. Las posiciones de otras celdas también difieren.
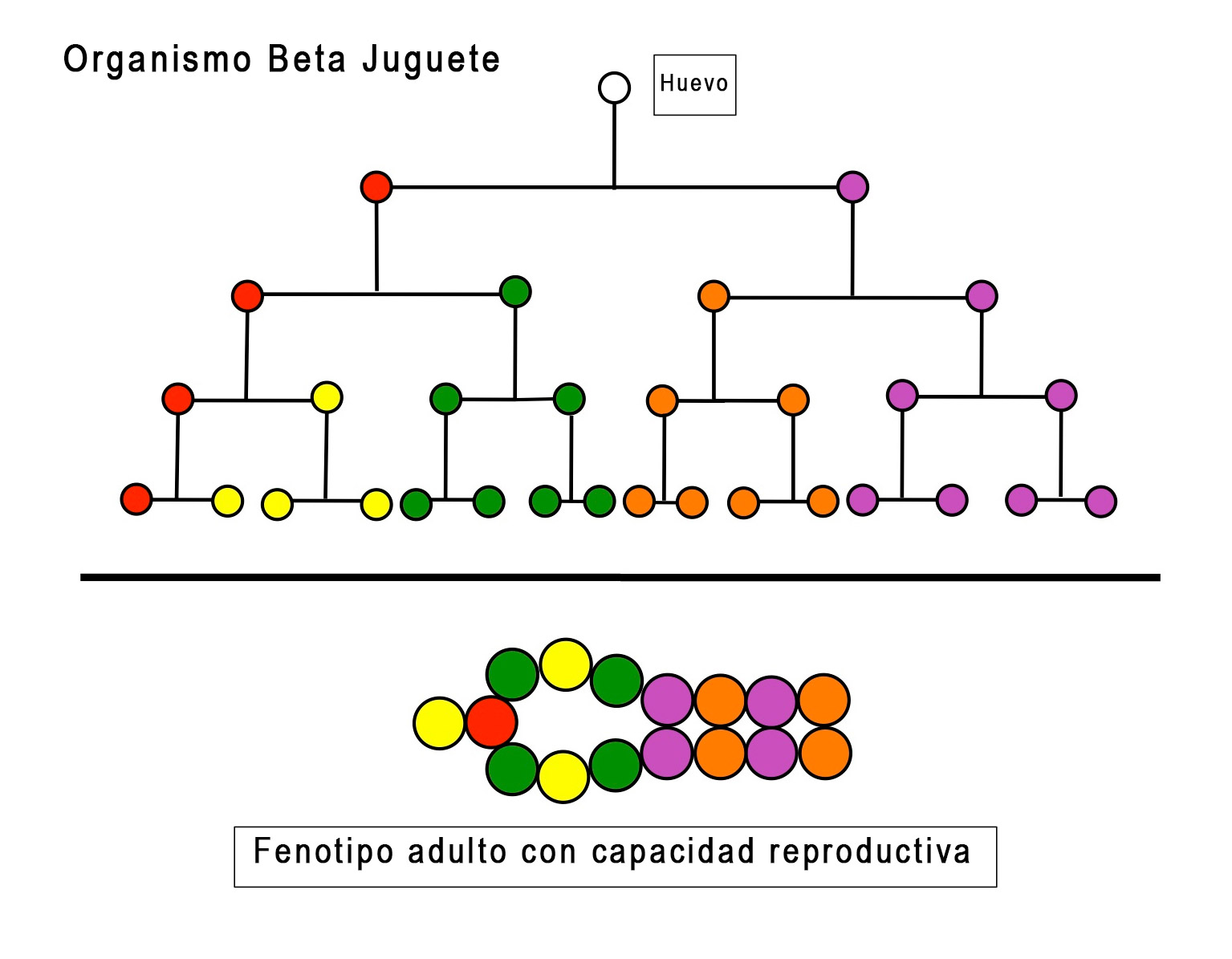
Figura 6
Si la capacidad reproductiva depende (al menos en parte) de una forma precisa, como lo hace para cualquier animal real, la medición de la profundidad ontogenética requerirá especificar las posiciones tridimensionales y las relaciones de las células o tejidos individuales. Esta información posicional no puede ser capturada ni por división celular (linajes) ni por tipo de celda, y proporcionar una métrica será enormemente más difícil.
«Entonces, ¿por qué preocuparse por una distancia que todavía no se puede medir?», Probablemente el lector esté preguntando. «¿Cuál es el punto de?»
4. La teoría de la evolución por selección natural exige que expliquemos la profundidad ontogenética
Es por eso. Como explicaré en la Parte II, el uso del proceso de selección natural en la explicación evolutiva impone exigencias evidentes estrictas al investigador. La evolución de las estructuras dirigidas a objetivos del desarrollo de metazoos de novo plantea desafíos que la selección natural─ya que el proceso carece de previsión─puede en principio ser incapaz de explicar.
Artículo publicado originalmente en inglés por Paul Nelson



{kind=link}